Blog Archives

We have good clostridia in the gut and some of them prevent allergies
21st March 2015
Clostridia: who are they ?
The clostridia or Clostridiales, with Clostridium and other related genera, are Gram-positive sporulating bacteria. They are obligate anaerobes, and belong to the taxonomic phylum Firmicutes. This phylum includes clostridia, the aerobic sporulating Bacillales (Bacillus, Listeria, Staphylococcus and others) and also the anaerobic aero-tolerant Lactobacillales (id est, lactic acid bacteria: Lactobacillus, Leuconostoc, Oenococcus, Pediococcus, Lactococcus, Streptococcus, etc.). All Firmicutes have regular shapes of rod or coccus, and they are the evolutionary branch of gram-positive bacteria with low G + C content in their DNA. The other branch of evolutionary bacteria are gram-positive Actinobacteria, of high G + C and irregular shapes, which include Streptomyces, Corynebacterium, Propionibacterium, and Bifidobacterium, among others.

Being anaerobes, the clostridia have a fermentative metabolism of both carbohydrates and amino acids, being primarily responsible for the anaerobic decomposition of proteins, known as putrefaction. They can live in many different habitats, but especially in soil and on decaying plant and animal material. As we will see below, they are also part of the human intestinal microbiota and of other vertebrates.
The best known clostridia are the bad ones (Figure 1): a) C. botulinum, which produces botulin, the botulism toxin, although nowadays has medical and cosmetic applications (Botox); b) C. perfringens, the agent of gangrene; c) C. tetani, which causes tetanus; and d) C. difficile, which is the cause of hospital diarrhea and some postantibiotics colitis.
Figure 1. The four more pathogen species of Clostridium. Image from http://www.tabletsmanual.com/wiki/read/botulism
Clostridia in gut microbiota
As I mentioned in a previous post (Bacteria in the gut …..) of this blog, we have a complex ecosystem in our gastrointestinal tract, and diverse depending on each person and age, with a total of 1014 microorganisms. Most of these are bacteria, besides some archaea methanogens (0.1%) and some eukaryotic (yeasts and filamentous fungi). When classical microbiological methods were carried out from samples of colon, isolates from some 400 microbial species were obtained, belonging especially to proteobacteria (including Enterobacteriaceae, such as E. coli), Firmicutes as Lactobacillus and some Clostridium, some Actinobacteria as Bifidobacterium, and also some Bacteroides. Among all these isolates, some have been recognized with positive effect on health and are used as probiotics, such as Lactobacillus and Bifidobacterium, which are considered GRAS (Generally Recognized As Safe).
But 10 years ago culture-independent molecular tools began to be used, by sequencing of ribosomal RNA genes, and they have revealed many more gut microorganisms, around 1000 species. As shown in Figure 2, taken from the good review of Rajilic-Stojanovic et al (2007), there are clearly two groups that have many more representatives than thought before: Bacteroides and Clostridiales.

Figure 2. Phylogenetic tree based on 16S rRNA gene sequences of various phylotypes found in the human gastrointestinal tract. The proportion of cultured or uncultured phylotypes for each group is represented by the colour from white (cultured) passing through grey to black (uncultured). For each phylogenetic group the number of different phylotypes is indicated (Rajilic-Stojanovic et al 2007)
In more recent studies related to diet such as Walker et al (2011) — a work done with faecal samples from volunteers –, population numbers of the various groups were estimated by quantitative PCR of 16S rRNA gene. The largest groups, with 30% each, were Bacteroides and clostridia. Among Clostridiales were included: Faecalibacterium prausnitzii (11%), Eubacterium rectale (7%) and Ruminococcus (6%). As we see the clostridial group includes many different genera besides the known Clostridium.
In fact, if we consider the population of each species present in the human gastrointestinal tract, the most abundant seems to be a clostridial: F. prausnitzii (Duncan et al 2013).
Benefits of some clostridia
These last years it has been discovered that clostridial genera of Faecalibacterium, Eubacterium, Roseburia and Anaerostipes (Duncan et al 2013) are those which contribute most to the production of short chain fatty acids (SCFA) in the colon. Clostridia ferment dietary carbohydrate that escape digestion producing SCFA, mainly acetate, propionate and butyrate, which are found in the stool (50-100 mM) and are absorbed in the intestine. Acetate is metabolized primarily by the peripheral tissues, propionate is gluconeogenic, and butyrate is the main energy source for the colonic epithelium. The SCFA become in total 10% of the energy obtained by the human host. Some of these clostridia as Eubacterium and Anaerostipes also use as a substrate the lactate produced by other bacteria such as Bifidobacterium and lactic acid bacteria, producing finally also the SCFA (Tiihonen et al 2010).
Clostridia of microbiota protect us against food allergen sensitization
This is the last found positive aspect of clostridia microbiota, that Stefka et al (2014) have shown in a recent excellent work. In administering allergens (“Ara h”) of peanut (Arachis hypogaea) to mice that had been treated with antibiotics or to mice without microbiota (Germ-free, sterile environment bred), these authors observed that there was a systemic allergic hyper reactivity with induction of specific immunoglobulins, id est., a sensitization.
In mice treated with antibiotics they observed a significant reduction in the number of bacterial microbiota (analysing the 16S rRNA gene) in the ileum and faeces, and also biodiversity was altered, so that the predominant Bacteroides and clostridia in normal conditions almost disappeared and instead lactobacilli were increased.
To view the role of these predominant groups in the microbiota, Stefka et al. colonized with Bacteroides and clostridia the gut of mice previously absent of microbiota. These animals are known as gnotobiotic, meaning animals where it is known exactly which types of microorganisms contain.
In this way, Stefka et al. have shown that selective colonization of gnotobiotic mice with clostridia confers protection against peanut allergens, which does not happen with Bacteroides. For colonization with clostridia, the authors used a spore suspension extracted from faecal samples of healthy mice and confirmed that the gene sequences of the extract corresponded to clostridial species.
So in effect, the mice colonized with clostridia had lower levels of allergen in the blood serum (Figure 3), had a lower content of immunoglobulins, there was no caecum inflammation, and body temperature was maintained. The mice treated with antibiotics which had presented the hyper allergic reaction when administered with antigens, also had a lower reaction when they were colonized with clostridia.
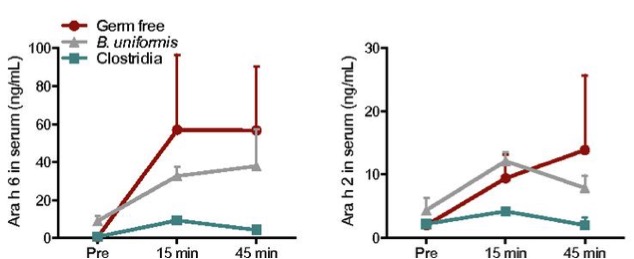
Figure 3. Levels of “Ara h” peanut allergen in serum after ingestion of peanuts in mice without microbiota (Germ-free), colonized with Bacteroides (B. uniformis) and colonized with clostridia. From Stefka et al (2014).
In addition, in this work, Stefka et al. have conducted a transcriptomic analysis with microarrays of the intestinal epithelium cells of mice and they have found that the genes producing the cytokine IL-22 are induced in animals colonized with clostridia, and that this cytokine reduces the allergen uptake by the epithelium and thus prevents its entry into the systemic circulation, contributing to the protection against hypersensitivity. All these mechanisms, reviewed by Cao et al (2014), can be seen in the diagram of Figure 4.
In conclusion, this study opens new perspectives to prevent food allergies by modulating the composition of the intestinal microbiota. So, adding these anti-inflammatory qualities to the production of butyrate and other SCFA, and the lactate consumption, we must start thinking about the use of clostridia for candidates as probiotics, in addition to the known Lactobacillus and Bifidobacterium.

Figure 4. Induction of clostridia on cytokine production by epithelial cells of the intestine, as well as the production of short chain fatty acids (SCFA) by clostridia (Cao et al 2014).
References
Cao S, Feehley TJ, Nagler CR (2014) The role of commensal bacteria in the regulation of sensitization to food allergens. FEBS Lett 588, 4258-4266
Duncan SH, Flint HJ (2013) Probiotics and prebiotics and health in ageing populations. Maturitas 75, 44-50
Rajilic-Stojanovic M, Smidt H, de Vos WM (2007) Diversity of the human gastrointestinal tract microbiota revisited. Environ Microbiol 9, 2125-2136
Rosen M (2014) Gut bacteria may prevent food allergies. Science News 186, 7, 4 oct 2014
Russell SL, et al. (2012) Early life antibiotic-driven changes in microbiota enhance susceptibility to allergic asthma. EMBO Rep 13(5):440–447
Stefka AT et al (2014) Commensal bacteria protect against food allergen sensitization. Proc Nat Acad Sci 111, 13145-13150
Tiihonen K, Ouwehand AC, Rautonen N (2010) Human intestinal microbiota and healthy aging. Ageing Research Reviews 9:107–16
Walker AW et al (2011) Dominant and diet-responsive groups of bacteria within the human colonic microbiota. The ISME J 5, 220-230
